
Bakterier
| Bakterier | |||
|---|---|---|---|
.jpg) Escherichia coli-celler forstørret 25 000 ganger
| |||
| Nomenklatur | |||
| Bacteria, Eubacteria | |||
| Populærnavn | |||
| bakterier, ekte bakterier | |||
| Hører til | |||
| liv | |||
| Økologi | |||
| Antall arter: | opp til 10 000[1][2] | ||
| Habitat: | overalt | ||
| Utbredelse: | hele jorden | ||
| Inndelt i | |||
| |||
Bakterier (gresk βακτήριον [bakterion] = «liten stav») er encellede mikroorganismer. De er typisk bare noen få mikrometer lange og kan ha mange forskjellige former, eksempler på disse fasongene er stavbakterier, kokker og spiriller. Studien av bakterier er en gren av mikrobiologien. Bakterier er allestedsnærværende på jorden og kan leve i alle slags miljøer. De har blitt påvist i jordsmonn, radioaktiv væske og varme kilder for å nevne noen.[3] Det er om lag 40 millioner bakterieceller i et gram jord og én million bakterieceller i en milliliter ferskt vann; i hele verden er det omtrent fem kvintillioner (5×1030) bakterier.[4] Bakterier spiller en vital rolle i resirkulering av næringsstoffer, og mange prosesser er avhengige av bakterier, slik som fiksering av nitrogen fra atmosfæren.
Flesteparten av disse bakteriene har imidlertid ikke blitt beskrevet, og kun halvparten av stamtreet har arter som kan bli dyrket frem i laboratoriet.[5] Det kan være mange flere bakterieceller enn menneskeceller i en menneskekropp, med høyest innhold på huden og i endetarmen.[6] Likevel er majoriteten av disse bakteriene uskadelige, men heller fordelaktig for «naturens husholdning» og de forskjellige livsprosessene.[7] Noen få bakterier er patogene for mennesker, eksempel på sykdommer som blir fremkalt av bakterier er kolera, syfilis, miltbrann og svartedauden. De mest fatale infeksjonssykdommene er de som hemmer åndedrettssystemet, med tuberkulose som alene dreper omtrent to millioner mennesker per år.[8]
Ukritisk bruk av antibiotika til å behandle bakterielle infeksjoner og i forskjellige jordbruksprosesser har gjort at antibiotikaresistens har blitt et stort problem, både i industrialiserte land og i utviklingsland. Restriktiv bruk av antibiotika bl.a. i Norge gjør at det her har vært et noe mindre problem. Forebygging, hygieniske tiltak og ikke-medikamentell behandling blir dermed viktigere, for eksempel vil mange bakterier hemmes eller drepes av sølv.
Bakterier er også viktig i industrien, de blir blant annet benyttet til å behandle avfallsvann, produksjon av ost og yoghurt, de blir også anvendt i produksjon av antibiotika og andre kjemikalier.[9]
Bakterier er prokaryoter. Ulikt dyreceller[10] og andre eukaryoter, inneholder bakteriecellene ingen cellekjerne og har sjelden membran-bundet organeller. Likeledes inkluderer begrepet bakterier tradisjonelt alle prokaryoter, den vitenskapelige nomenklaturen etter oppdagelsen av prokaryotisk liv består av to forskjellige typer av organismer som utviklet seg uavhengig av hverandre fra samme stamfar. Disse evolusjonære områdene blir kalt bakterier og archaea.[11]
Bakterienes plassering[rediger | rediger kilde]
Bakterier regnes innenfor tradisjonell systematikk som et av «rikene» av liv innenfor biologien. Man deler ofte bakteriene inn i ekte bakterier og arkebakterier. Ofte opererer man med følgende evolusjonære hovedgrupper eller «riker» i den taksonomiske systematikken:[12]
- Prokaryoter:
- Bakterier (Bacteria, Eubacteria)
- Arkebakterier (Archaea)[13]
- Eukaryoter:
- Protister (Protista)
- Planter (Plantae)
- Sopp (Fungi)
- Flercellede dyr (Metazoa)
Dette systemet med «riker» er fortsatt omstridt. Den svenske Nationalnyckeln till Sveriges flora och fauna[14] opererer med 5 riker, mens en gruppe europeiske biologer[15] observerer 3 basale grupper hvor bakterier og arkebakterier utgjør to, og den siste gruppen av eukaryoter siden gir opphav til protister, planter, sopp og dyr. Systematikkens «riker» representerer et forsøk på å forene behovet for oversikt med behovet for å dele livsformene opp i mest mulig naturlige grupper.
Bakteriologiens historie[rediger | rediger kilde]

Nederlenderen Anton van Leeuwenhoek var den som først oppdaget bakteriene (1674). Louis Pasteur og Robert Koch forsket videre på bakterier, og grunnla en egen bakteriologisk vitenskap.
. . . mitt arbeid, som jeg har holdt på med i lengre tid, ble ikke forfulgt for å nå den anerkjennelse jeg nå nyter men hovedsakelig som et begjær etter kunnskap, som jeg merker jeg har mer av enn de fleste andre menn. Og følgelig, når jeg har funnet ut noe bemerkelsesverdig, har jeg sett det som min plikt å nedtegne på papir min oppdagelse, så alle oppfinnsomme mennesker kan informeres om dette.
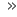 Anton van Leeuwenhoek, 1716
Anton van Leeuwenhoek, 1716
Robert Koch var den første til å dyrke bakterier på et fast medium. Koch benyttet gelatin som løsemiddel for næringsløsningene han brukte til å dyrke frem patogene (skadelige) bakterier og utviklet med det en metode for å dyrke frem bakterier på en fast plate som var fri for forurensing. Han la et sylteglass eller et glass over platen for å unngå kontaminasjon.
Gelatin var fantastisk for å dyrke frem bakterier i et kulturmedium. Gelatin hadde likevel sine ulemper, da gelatin ikke er i fast form ved kroppstemperatur (37 °C), den optimale temperaturen for dyrking av patogene bakterier for mennesker. Det måtte finnes et mer allsidig vekstmedium, og dette viste seg å være agar.
Agar er et polysakkarid som utvinnes fra røde alger.[16] Walter Hesse var den første til å bruke agar som vekstmedium. Ideen om å bruke agar som vekstmedium kom fra Hesses kone, Fannie. Fannie Hesse brukte agar for å tilberede syltetøy og når Walter prøvde det ut som vekstmedium fungerte det utmerket. Walter skrev til Robert Koch om oppdagelsen og Koch bestemte seg raskt for å bruke agar i sine egne studier.
I 1887 publiserte Julius Richard Petri en artikkel som omhandlet en modifikasjon av Kochs plate. Petris innblanding viste seg å være av stor betydning. Den nye metoden gjorde det mulig å sterilisere separat fra mediet og flytte kolonier fra en mindre plate til for eksempel to større. Den originale ideen om platen er ikke forandret den dag i dag og platen har blitt oppkalt etter Petri. En Petri-plate er laget av enten glass som blir sterilisert av varme eller laget av plast som blir sterilisert av etylenoksid. Denne nye platen hadde stor betydning for den videre forskningen på mikroorganismer.
Koch oppdaget at forskjellige kolonier som hadde utviklet seg på en kontaminert plate hadde forskjellig form (farge, morfologi, størrelse). Disse forskjellene i koloniform gjorde at man klarte å skille ulike bakterier fra hverandre på platen. Man klarte også å skille forskjellige celler i mikroskopet, og ved hvilke temperaturer og næringsstoffer de trives i.
Robert Koch
Kochs oppdagelser av kulturmedium og iver etter å dyrke rene bakteriekulturer gikk langt utover hva man kunne forvente på den tiden. Verden står i stor takk til Robert Koch og hans medhjelpere for deres innsats innenfor bakteriologi og mikrobiologi.
Opphav og tidlig evolusjon[rediger | rediger kilde]
Utdypende artikkel: Evolusjon
Stamfaren til nåtidens bakterier var encellede mikroorganismer som var den første formen for liv på vår jord for omtrent fire milliarder år siden.[17] For ca. 3 milliarder år siden var alle organismer mikroskopiske, og bakterier og archaea var den dominerende formen for liv.[18] Det har blitt funnet fossiler av bakterier som stromatoliter, men mangelen på morfologi gir oss liten nytte for å studere deres evolusjon eller å datere deres tidsløp. Det er mulig å bruke genteknologi til å rekonstruere deres fylogenetikk, og disse undersøkelsene tyder på at bakteriene har et opphav fra to ulike organismer, erkebakterien og eubakterien.[19] Den siste stamfaren til bakterier og archaea var sannsynligvis en hypertermofil som levde rundt 2,5-3,2 milliarder år siden.
Eldste bevis for liv[rediger | rediger kilde]
Det eldste beviset man har for liv på jorden er nylig avdekte mikrofossiler funnet i Nunavik i Quebec, Canada. Funnet (som ble publisert 1. mars 2017) dateres cirka 3 770 millioner år tilbake i tid.[20] Før denne oppdagelsen var det eldste beviset mikrofossiler funnet i Vest-Australia, datert cirka 3 460 millioner år tilbake.[20]
Det var en internasjonal forskergruppe, under ledelse av University College London, som gjorde funnet. Det besto av en fossil bakterie som levde på jern, og funnet ble gjort i et lag med kvarts. Gruppen av forskere besto av folk fra flere land og organisasjoner, blant annet fra Norges geologiske undersøkelser.[20]
Morfologi og stamtre[rediger | rediger kilde]

Bakterienes systematiske inndeling er fortsatt debattert. Dette kommer av at «klassiske» metoder for slektskapsrekonstruksjon, som tar utgangspunkt i morfologi, har få holdepunkt å gå etter, fordi bakterieceller er så enkelt bygd.
Molekylære metoder har derfor revolusjonert bakterie-systematikken, men gir sprikende resultater. Grunnene er at bakterier kan utveksle gener mellom ulike bakteriearter, til og med mellom nokså fjernt beslektede. Blant kandidatene til den mest basale grenen i bakterienes stamtre er for eksempel både Planctomycetes, Thermotoga maritima og Aquificae. Delgruppene som er nevnt i boksen øverst til høyre, er basert på den nyeste kunnskapen (per 2004). Man vil likevel måtte forvente en del endringer i fremtiden, både grunnet bedre fylogenetiske analysemetoder og fordi nye arter vil bli beskrevet.
Andre vanlige inndelinger er etter utseende, levevis eller celleveggens bygning, men de færreste av disse peker ut naturlige slektskapsgrupper.
Inndeling etter utseende[rediger | rediger kilde]
På samme måte som ulike planter og dyr har forskjellig utseende (morfologi) har bakterier ulikt utseende, både på makroskopisk og mikroskopisk nivå. For bakteriologer/mikrobiologer er det viktig å kunne skille de ulike bakterieartene. Dette kan gjøres ved å undersøke deres koloniutseende på forskjellige vekstmedia for så å definere koloniens morfologi.
I tillegg til dette har ulike bakterier forskjellig mikroskopisk utseende og deles da gjerne inn i klasser i henhold til fremtoning. En bakterie som er kuleformet eller sirkulær i utseende kalles kokk. Kokker kan igjen grupperes avhengig av om de vokser i par, kjeder eller fire og fire, eller i klaser. En avlang eller sylinderformet bakterie kalles stav. Det finnes også mer sjeldne morfologiske typer spiralformede og filamentære bakterier.
- kokker (kuleformede)
- staver (stavformede; jf. bakterienes navn)
- vibrioner (kommaformede)
- spirochæter (skrueformede)
- spiriller (spiralformede)
Inndeling etter levevis[rediger | rediger kilde]
- saprofytter – som lever av dødt materiale
- parasitter – som snylter på andre levende organismer
- symbionter – som lever i et gjensidig nytteforhold med andre organismer
- autotrofe – som er selvforsynt med energi vha. kjemo- eller fotosyntese
Fordelt på utseende er det kun tre typer som er patogene (dvs. parasittiske):
- Staver kan gi for eksempel difteri, kikhoste, spedalskhet, stivkrampe, tuberkulose og tyfoidfeber
- Kokker kan gi for eksempel blodforgiftning, hjernehinnebetennelse, lungebetennelse og skarlagensfeber
- Spirochæter kan gi for eksempel syfilis, borrelia og flekktyfus
Nanobakterier[rediger | rediger kilde]
Utdypende artikkel: Nanober
Noen mikrobiologer har foreslått en type veldig små bakterier i naturen, celler som blir kalt for nanobakterier[21] på grunnlag av svært små bakterieliknende strukturer som kan dukke opp i stein. Størrelsen på slike nanobakterier er antatt til å være under 0,1 µm for kokkeformet strukturer. Skeptikere mener nanobakterier er enkle mekanismer av kjemiske eller geokjemiske reaksjoner av ikke-levende materiale. De peker på at de minste bakteriene vi kjenner til er betydelig større enn disse antatte nanobakteriene. Et annet viktig argument er at de er for små til å inneholde essensielle biomolekyler for liv. Det er høyst usannsynlig at disse molekylene kan eksistere i et volum av 0,1 µm eller mindre. Spørsmålet om nanobakterier er liv eller geologi er ikke klarlagt, det samme gjelder spørsmålet om den nedre grensen for størrelsen på levende organismer. Blir nanobakterier påvist vil de være den minste formen for liv vi kjenner til.[22]
Cellestruktur[rediger | rediger kilde]


2. Fettsyrer
3. Polysakkarider (arabinogalactan)
4. Peptidoglycan
5. Plasma membran
6. Lipoarabinomannan (LAM)
7. Fosfatidylinositol mannoside
8. Cellevegg skjelett
Bakterier kan inndeles i to hovedgrupper, Gram-positive (G+) og Gram-negative (G-). Dette kommer originalt fra en spesiell metode som benevnes gramfarging (Gram stain).[23] Når denne metoden benyttes får cellene ulik farge avhengig om det er G+ eller G-. Det er forskjell i oppbygningen i celleveggen som er årsaken til dette. G- cellenes vegger har en komplisert oppbygging i flere lag, mens mens G+ har ett enkelt tykt lag av peptidoglycan.[24][25]
Ekstracellulær struktur[rediger | rediger kilde]
Konsentrasjonen av oppløste stoffer i bakterieceller er høy, dette gjør at det blir et høyt trykk inne i cellen. En typisk Escherichia coli har et trykk på 2 atm.[24] Det er celleveggens jobb å motstå dette trykket. Prokaryoters cellevegg består hovedsakelig av peptidoglycan[26] og er vanskelig å se i et lysmikroskop, men kan bli sett i et elektronmikroskop.
Celleveggen ligger utenfor den cytoplasmatiske membranen. Stivheten og styrken til celleveggen kommer hovedsakelig av innholdet i peptidoglycan. Den er også hovedårsak til hvilken form bakterier får.[26] Peptidoglycan blir også kalt for murein i eldre kilder.
Peptidoglykan[rediger | rediger kilde]
Peptidoglykan finnes både hos Gram-positive og Gram-negative bakterier, men hos Gram-negative er det flere lag av andre stoffer utenpå dette stive laget (se bilde). Peptidoglycan består av to sukkerderivater, N-acetylglukosamin og N-acetylmuraminsyre og en liten gruppe av aminosyrer.[25] Det finnes over 100 variasjoner av oppbygningen av peptidoglykan og de mest vanlige aminosyrene er: L-alanin, D-alanin, D-glutaminsyre og lysin eller diaminopimelsyre (DAP). Disse bestanddelene er koblet sammen i en repetert struktur som kalles glykan tetrapeptide.[24]
Strukturen til peptidoglykan er koblet sammen i flak hvor glykankjeder som blir dannet fra sukker blir koblet sammen av peptid-kryssbindinger som er dannet av aminosyrer.[25] Glykogenet som binder sammen sukkeret i kjedene er veldig sterkt, men disse kjedene alene er ikke nok til å gi ubøyelighet/stivhet i alle retninger. Peptidoglykanets fulle styrken kommer først når det blir kryssbundett av aminosyrene. Kryssbindeingen er karakteristisk for de forskjellige bakteriene. Jo mer stiv en bakterie er jo mer kompleks krysslinking.
I Gram-negative bakterier blir dette gjort ved direkte linking av aminogruppen diaminopimelsyre til karboksylgruppen i D-alanin. Gram-positive krysbindes av en peptidbru som varierer hos forskjellige bakteriegrupper. Hos gule stafylokokker (Staphylococcus aureus), en godt studert G+ bakterie, er peptidbruen koblet sammen av fem aminosyrer.
Peptidoglykan er bare påvist i bakterier. Det har ikke blitt påvist i Archaea (arkebakterier eller urbakterier) eller eukaryote organismer (Eukarya). Likevel har ikke alle bakterier DAP i sitt peptidoglykan. Denne aminosyren er påvist i alle Gram-negative og i noen få Gram-positive bakterier. Kokkeformede G+ bakterier har lysin i stedet for DAP, og noen få G+ har andre aminosyrer. En annen bemerkelsesverdig ting med celleveggen er at alle aminosyrene har D-konfigurasjon (se stereoisomeri). Aminosyrer som er byggesteiner for proteiner har alltid L-konfigurasjon hos mennesker.[24]
Yttermembran til Gram-negative[rediger | rediger kilde]
Utenom peptidoglycan består Gram-negative celler et lag utenfor som består av lipopolysakkarid.[23] Dette laget er et effektivt bilag, men består ikke utelukkende av fosforlipider, som den cytoplasmatiske membranen gjør. I stedet er den oppbygd av polysakkarider og proteiner. Lipid og polysakkaridet er inngående lenket i det ytre laget til den ytre membran, ved å danne spesifikke lipopolysakkaridstrukturer. På grunn av tilstedeværelsen av lipopolysakkarid, blir dette laget kalt for lipopolysakkarid laget, eller LPS. Et annet utbredt begrep er «den ytre membran».
Hovedfunksjonen til den ytre membran er strukturen, men en viktig biologisk funksjon er at den ofte er giftig (endotoksisk) for mennesker og pattedyr. Av denne grunnen er Gram-negative bakterier patogene ovenfor mennesker og andre pattedyr.[27] Årsaken til dette er lipopolysakkaridet som består av en bestanddel Lipid A. Noen eksempler på Gram-negative bakterier som er patogene for mennesker er Salmonella, Shigella og Escherichia m.f.
Intracellulær struktur[rediger | rediger kilde]

Bakteriecellene er omgitt av en lipid membran, eller cellemembran som omringer innholdet av cellen og fungerer som en barriere. Næringsstoffer og andre stoffer som er viktige for cellen kan gå igjennom og på samme måte kan avfallsstoffer og andre celleprodukter gå ut. Ettersom bakterier er prokaryoter har de ikke membran-bundete organeller i cytoplasma og inneholder bare noen få intracellulære strukturer. Bakterier mangler mitokondrier, cellekjerne, kloroplast og andre organeller som eukaryotiske celler har, som for eksempel endoplasmatisk retikulum, og golgiapparatet.[28]
Mange viktige biokjemiske reaksjoner, slik som energiutvikling, skjer ved at konsentrasjonen stiger tvers over membranen og lager en potensialforandring som et batteri.
DNA er ikke organisert i en cellekjerne, men cellen har i stedet et enkelt, ringformet kromosom. Ofte finner man i tillegg såkalte plasmider, dvs. DNA som ikke er organisert i kromosomer. Som alle levende organismer inneholder bakterier ribosomer for produksjon av proteiner, men strukturen til bakterie ribosomene er forskjellig fra eukaryotiske- og archaea celler.[29]
Endosporer[rediger | rediger kilde]
Utdypende artikkel: Endosporer
Bestemte typer bakterier produserer i dårlige tider spesielle strukturer som kalles endosporer (endo: inne) i cellene (se bilde til høyre). Prosessen kalles sporulation. Sporer er celler som er veldig resistent mot varme. De er også resistente ovenfor sterke kjemikalier.[30] I jord er det rikelig med bakterier som danner sporer, og en hvilken som helst jordprøve vil inneholde endosporer. Slektene staver/bacillus og Clostridium er de best studerte artene som danner sporer.[31]
Funnet av endosporer var en viktig oppdagelse fordi det er viktig å vite om varmeresistente former for bakterier ved sterilisering av utstyr. Det er ikke bare bakterier som danner sporer, men de bakterielle sporene er unik i at de tåler høye temperaturer. Foruten å tåle varme, tåler endosporene tørking, radioaktivitet, syrer og andre sterke kjemikalier. De kan være uvirksomme sporer i ekstremt lang tid.[30]
Strukturen til sporene er overveldende forskjellig fra selve cellen. Denne er mye mer kompleks i det at de har flere lag som man ikke finner i cellen. En kjemisk substans som er karakteristisk for endosporer er dipicolinsyre som ligger i "kjernen" av sporen. Dette finnes ikke i bakteriecellene. Dette stoffet har blitt funnet i alle endosporene som har blitt undersøkt. Sporene har også en høy konsentrasjon av kalsium ioner som er kombinert med dipicolinsyren. Denne kalsium-dipicolinsyren står for omtrent 10 % av tørrvekten til sporene.[24]
Forskjeller fra endosporer og vegetabilske celler[rediger | rediger kilde]
| Karakteristikk | Vegetabilsk celle | Endosporer |
|---|---|---|
| Struktur | Typisk gram-positiv celle og noen få gram-negative celler | Tykt lag, spore kåpe, exosporium |
| Kalsium innhold | Lav | Høy |
| Dipicolinsyre | Inneholder ikke | Inneholder |
| Enzymaktivitet | Høy | Lav |
| Metabolisme (O2 opptak) | Høy | Lav eller ingen |
| Syntetisering av makromolekyler | Til stede | Ingen |
| mRNA | Til stede | Lav eller ingen |
| Resistens mot varme | Lav | Høy |
| Resistens mot radioaktivitet | Lav | Høy |
| Resistens mot kjemikalier (f.eks H2O2) og syrer | Lav | Høy |
| Mulighet for farging | Mulig | Kun ved spesielle teknikker |
| Innhold av vann | Høy, 70–80 % | Lav, 10–25 % i kjernen |
| pH i cytoplasma | Omtrent pH 7 | Omtrent pH 5,5–6 (kjerne) |
- (Tabellen er hentet fra Michael T. Madigan, John M. Martinko, Jack Parker (2003) Brock Biology of Microorganisms, Tenth Edition[24])
Metabolisme[rediger | rediger kilde]
Til forskjell fra høyere organismer, har bakterier en stor variasjon av metabolismer.[32] En nøkkelfunksjon til cellene er deres mulighet til å omsette kjemiske reaksjoner og organisere molekyler til spesifikke strukturer. Bakteriene er fleksible ved at de kan skifte metabolisme avhengig av omgivelsene.[33] Hovedproduktet av dette er vekst (replikasjon). Før dette kan skje må en rekke kjemiske reaksjoner i gang som samlet blir kalt for metabolisme. Metaboliske reaksjoner er enten energifrigivende (katabolisk reaksjon) eller energikrevende (anabolisk reaksjon).[34]
Cellene består hovedsakelig av makromolekyler (som igjen består av monomerer) og vann. Cellenes oppgave er å skaffe disse næringsstoffene til å lage monomerer. Forskjellige organismer trenger forskjellige sett med næringsstoffer og de får disse molekylene i en eller annen form. Ikke alle næringsstoffene trengs i lik grad, de som trengs i størst skala blir kalt «makronæringsstoffer» og de i minst skala «mikronæringsstoffer».[35]
Mange prokaryoter trenger organiske komponenter som deres kilde til karbon. Studier viser at bakteriene kan benytte ulike typer organiske molekyler som kilde til nytt cellemateriale. Aminosyrer, fettsyrer, organiske syrer, sukker, nitrogen baser, aromatiske komponenter og et uendelig hav av andre molekyler som blir brukt som næringsstoff av ulike bakterier.[24] Noen typer bakterier er autostrofer som betyr at de klarer å bygge organiske komponenter fra karbondioksid (CO2)[33]. Energien får de fra enten lys eller uorganiske molekyler. Karbon er det viktigste grunnstoffet til bakterier, og en typisk bakteriecelle består av om lag 50 % karbon (tørrvekt).
Etter karbon er nitrogen det viktigste næringsstoffet for bakterier. En typisk bakteriecelle består av ca. 12 % nitrogen (tørrvekt). Nitrogen er et viktig element i proteiner, nukleinsyrer og en rekke andre komponenter i cellen. Grunnstoffet finnes i både organisk og uorganisk form i naturen. Hovedmassen av nitrogen finnes i uorganisk form som ammoniakk, nitrat eller nitrogengass. De fleste bakteriene klarer å nære seg av ammoniakk alene, men mange andre kan også bruke nitrat. En spesiell type nitrogenfikserende bakterier kan livnære seg på nitrogengass.[36]
Fosfor forekommer i naturen i form av organisk- og uorganiske forbindelser. Dette grunnstoffet er viktig ved syntetisering av nukleinsyre og fosforlipider. Svovel er viktig for strukturelle roller i aminosyrene cystein og metionin. Det blir også brukt i forskjellige vitaminer og koenzym A. Hovedsakelig får cellene svovel fra sulfat (SO42-) og sulfid (HS-).[37] Kalium er nødvendig for alle typer organismer. En stor variasjon av enzymer og spesielt de som deltar i proteinsyntesen trenger kalium. Magnesium fungerer som en stabilisator for ribosomer, cellemembran og nukleinsyrer. Det er og nødvendig for aktiviteten til enzymer. Kalsium er ikke et nødvendig næringsstoff for celler, men spiller en viktig rolle i varmestabilisering i endosporer.
Vekst og reproduksjon[rediger | rediger kilde]

Bakteriecellene har evnen til å kopiere seg selv. Ved denne syntetiseringen skjer det rundt 2000 forskjellige kjemiske reaksjoner av ulike slag. Noen av reaksjonene involverer transport av energi. Andre av disse reaksjonene er omforming av energi og andre involverer biosyntetisering av små molekyler (byggesteiner for makromolekyler). Før cellen kan dele seg må nytt DNA syntetiseres, etter dette må andre cellekomponenter som cytoplasmatisk membran, flageller, ribosomer etc. lages.
I de fleste prokaryoter fortsetter veksten av en individuell celle til to nye celler har blitt dannet. Denne prosessen kalles binær fisjon (binær fordi én celle blir til to). I en vekstkultur av stavformede bakterier, som for eksempel Escherichia coli vokser staven til det dobbelte av sin egen lengde. Etter dette deler den seg til to datterceller. Denne prosessen blir kalt for septum og skjer ved at den cytoplasmatiske membranen blir dratt innover til midten i cellen slik at den deles i to. Under vektsyklusen øker alle cellekomponentene slik at dattercellene får egne sett av kromosomer, makromolekyler, monomerer og uorganiske ioner for å kunne leve selvstendig.
Tiden det tar før en celle deler seg er høyst variabel og avhenger av en rekke forhold, både næringsstoffer, temperatur og genetiske faktorer. Når E.coli har gunstige forhold klarer den å dele seg på ca. 20 minutter, noen få bakterier klarer å dele seg raskere enn dette, men de fleste deler seg saktere.
En rekke proteiner har blitt påvist å være viktige i celledelingen. Disse kalles Fts proteiner. (Fts står for filamentous temperature sensitive) FtsZ er et nøkkelprotein i gruppen og er godt undersøkt i E.coli og en rekke andre bakterier. Fts proteiner finnes i alle prokaryoter inkludert arkebakterier. Ftp-Z type proteiner har også blitt funnet i mitokondrier og kloroplast. Proteinet har og en struktur som er lik tubulin som er et viktig protein i celledelingen hos eukaryoter. Disse funnene og likhetene tyder på at eukaryoter kan stamme fra prokaryoter[24]
Ftps proteiner påvirker hverandre til å danne et deleapparat som kalles divisome. Dannelsen av divisomet begynner med å sette fast molekyler av FtsZ i en ring rundt cellesylinderen i midten av cellen. Her blir cellen delt i to. FtsZ molekylene polymerer[38] og danner en intakt ring og ringen blir festet av andre fts proteiner. DNA replikasjon skjer før FtsZ ringen blir dannet og fungerer som et signal for dannelse av denne ringen.
Fordelen av å være liten[rediger | rediger kilde]
Små celler har bedre tilgang til næringsstoffer enn store. Det ser vi ved at alle næringsstoffer cellen trenger må transporteres gjennom cellemembranen. Arealet av denne er proporsjonal med kvadratet av cellens diameter. Samtidig er cellens behov for næringsstoffer bestemt av volumet til cellen, som er proporsjonalt med diameteren i tredje potens. Forholdet mellom overflaten og volumet er altså omvendt proporsjonalt med cellens diameter.[39]
Genetikk[rediger | rediger kilde]
De fleste bakteriene har bare ett kromosom, som er sirkulært, dvs at DNAet ligger i en lukket sløyfe istedenfor en åpen «lineær» streng som hos mennesket. Dette varierer i størrelse fra 160 000 basepar i bakterien Candidatus Carsonella ruddii,[40] til 12 200 000 basepar i Sorangium cellulosum.[41] Noen bakterier, som Borrelia bakterien Borrelia burgdorferi, som er årsaken til borreliose (lyme sykdom),[42] inneholder ett lineært kromosom.[43] Bakterier kan også inneholde plasmider, dette er små ekstra-kromosome DNA som kan replikere seg uavhengig av resten av DNAet i cellen. Det er kjent at disse kan overføres mellom bakterier i såkalt horisontal genoverføring. Hvis plasmidene inneholder gener for for eksempel antibiotikaresistens, er det en fare for at disse genene kan spres til andre arter av bakterier.[44] Andre typer av bakteriell DNA er integrerte virus (bakteriefager). Mange typer av bakteriefager finnes, noen enkle smitter og ødelegger bakterien, mens andre infiserer bakteriens kromosom. En bakteriefag kan inneholde gener som kan tilføres til en bakteries fenotype. Et eksempel er evolusjonen til Escherichia coli O157:H7 og Clostridium botulinum. Giftgenene hos disse stammer fra en integrert bakteriefage som gjorde en ufarlig forfader-bakterie til en dødelig en.[45]
Bakterier har ikke noe kjønn, de arver identiske kopier av sine foreldres gener (dvs. de er klonet). Likevel kan bakterier ha en evolusjon, gjennom naturlig utvalg og mutasjoner, samt horisontal genoverføring. Mutasjoner varierer fra art til art og til og med fra kloner innenfor en enkelt art av bakteriene.[46] Mutasjoner kommer enten av tilfeldig forandringer i DNAet ved deling eller «stress-styrt mutasjon», hvor genene er innblandet i en særskilt vekst-begrensning som øker sjansene for mutasjoner.[47]
Overføring av DNA mellom bakterier kan skje på tre ulike måter. Først, kan bakterier ta opp gener fra miljøet rundt, i en prosess som kalles transformation (norsk: omforming). Gener kan også bli overført ved hjelp av virus, såkalt transduction og når integreringen av en bakteriefag introduserer fremmed DNA til kromosomene. Den tredje og siste måten er genoverføring ved bakteriell konjugasjon[48], hvor DNA blir overført med direkte kontakt. Denne genoverføringen fra en bakterie eller fra miljøet rundt blir kalt for horisontal genoverføring og kan være vanlig under naturlige omstendigheter.[49]
Bevegelse[rediger | rediger kilde]

Mange prokaryotiske celler kan bevege seg. Dette gjøres ved hjelp av en spesiell struktur i eller utenfor cellen. Bestemte celler kan bevege seg på faste overflater ved gliding og bestemte vannlevende mikroorganismer kan bevege seg i vann ved hjelp av gass-blærer. Den vanligste måten å bevege seg på er ved hjelp av flageller. Bevegelse gjør at cellen kan nå andre regioner i miljøet sitt. I kampen for å overleve er dette viktig ved at bakterien kan forflytte seg til et nytt område med nye ressurser og muligheter.[50] Bakteriene beveger seg ved at motoren til flagellen roterer og kan nå flere 100 Hz, slik at bakteriecellene kan bevege seg flere kroppslengder per sekund.[51] Dette er en ressurskrevende prosess.
Bakterieflageller[rediger | rediger kilde]
- Utdypende artikkel: Flagell
Bakterieflagellen er en lang og tynn tilføyelse til bakteriecellen. Den er så tynn (ca. 20 nm) at man ikke kan se den direkte i lysmikroskop. Ved å tilsette en spesiell farge klarer man å øke diameteren til flagellen slik at den kan sees i lysmikroskop.
Flageller er arrangert forskjellig avhengig av type bakterie. I polar flagellisjon er flageller bundet til en eller begge endelsene (bilde: figur A og C) til bakterien. Av og til kan flagellene bli arrangert som en «gruppe» på enden til cellen (bilde: figur B), dette kalles lofotrikk (lopho betyr «gruppe», thrix «hår»). Peritrikk flagellisjon[52] (peri betyr «rundt») er flagellene festet forskjellige steder rundt bakterien (bilde: figur D). Type flagell brukes i klassifikasjonen av bakterier.
Flagellstruktur[rediger | rediger kilde]
Bakterieflagellene er en av de mest komplekse organellene i bakterien. Prosessen av transkripsjon og montering er ikke fullstendig forstått.[51]
Flagellen er ikke rett, men spiralformet. Når de ligger flatt vises en konstant avstand fra omkringliggende kurver som kalles bølgelengde. Denne bølgelengden er konstant for en bestemt organisme. Fiberet i flagellen består av subenheter av et protein kalt for flagellin.[53][54] Basisstrukturen til flagellene varierer lite fra de ulike typene av bakterier.
Fundamentet er forskjellig fra fiberet i flagellen. Det er et bredere område på fundamentet som kalles «kroken». Kroken består av et enkelt protein og funksjonen er å koble sammen «motoren» til fiberet. Motoren er festet til den cytoplasmatiske membran og celleveggen, motoren består av en liten sentral stang som passerer gjennom et system av ringer. I Gram-negative bakterier er en ytre ring forankret i lipopolysakkaridlaget og en annen i peptidoglycanlaget av celleveggen. En annen ring er og lokalisert inne i det cytoplasmatiske membranen. Gram-positive bakterier (som mangler lipopolysakkaridlaget) inneholder bare et par ringer som er festet i den cytoplasmatiske membranen. Der hvor disse ringene omkranser den indre ringen er et par av proteiner som kalles Mot. Disse proteinene fungerer som vridningsmoment for motoren og gjør at fiberet roterer. Et siste protein som kalles Fli virker som en avbryter og reverserer rotasjonen av flagellen fra intracelluare signaler.
Flageller fra en unik gruppe av bakterier, spirochaetes, er funnet mellom to membraner i det periplasmiske rommet.[55]
Samspill med andre organismer[rediger | rediger kilde]
Til tross for sin tilsynelatende enkelhet kan bakterier ha komplekse samspill med andre organismer. Denne symbiotiske foreningen kan bli delt inn i parasittisme, mutualisme og kommensialisme På grunn av deres lille størrelse, lever bakteriene på samme måte på dyr og planter som på hvilken som helst overflate. Likevel kan deres vekst påvirkes av varme og svette. Store populasjoner av disse organismene på mennesker forårsaker svettelukt.
Mutualisme[rediger | rediger kilde]

Bestemte bakterier danner nære romlige bånd mellom hverandre, som er absolutt nødvendig for deres overlevelse. Ett eksempel på et slikt mutualistisk bånd kalles «interart hydrogen transport».[56] Dette oppstår mellom klynger med anaerobe bakterier som konsumerer organiske syrer som smørsyre og propansyre og danner hydrogen, og de metanfikserende Aechaea som konsumerer hydrogen.[57] De anaerobe bakteriene greier ikke å forbruke de organiske syrene på grunn av dette reaksjonen danner hydrogen som hoper seg opp i miljøet rundt. Hydrogen-konsumerende Archaea holder konsentrasjonen av hydrogen lav og slik klarer bakteriene å leve og å dele seg.
I den delen av jordsmonnet som kalles rhizosfæren, (en sone i jordvolumet som blir påvirket av planterotens tilstedeværelse) holder det til nitrogen-fikserende bakterier som konverterer nitrogen til nitrogenholdige komponenter.[58] Dette tilfører mange planter en lett absorberbar form av nitrogen som de ikke klarer å lage selv. Mange andre bakterier lever i symbiose med mennesket og andre organismer. For eksempel lever over 1 000 bakteriearter i normalfloraen i tarmen og kan bidra til immunitet, syntetisering av vitaminer som folsyre, vitamin K og biotin, konvertering av melkeproteiner til melkesyre, og til å gjære komplekse ufordøybare karbohydrater.[59][60][61] Tilstedeværelsen av denne normalfloraen forhindrer også vekst av potensielle patogene bakterier (hovedsakelig gjennom «å ta opp plass» i vevet) og disse gunstige bakteriene blir solgt som probiotika, kosttilskudd av bakterier.[62][63]
Patogener[rediger | rediger kilde]

Hvis en bakterie har et parasittisk samspill med andre organismer, klassifiseres de som patogener. Patogene bakterier er en stor årsak til menneskedød og sykdom og forårsaker infeksjoner slik som tetanus, tyfoidfeber, difteri, syfilis, kolera, matforgiftning, spedalskhet og tuberkulose. En patogenisk årsak til en kjent sykdom kan bli oppdaget mange år etter kontaminering av bakterien, et eksempel på en slik hendelse er Helicobacter pylori og magesår. Sykdommer forårsaket av bakterier er også viktig innen jordbruk med bakterier som årsak av pærebrann, wilt (en sykdom som får planter til å visne), bladflekksyke, og Johnes sykdom, brystkjertel infeksjon, salmonella og miltbrann i husholdningsdyr.
Hver art av patogener har sin egen karakteristikke spektrum av interaksjoner hos sin vert. Noen organismer, som Staphylococcus eller Streptococcus, kan gi årsak til hudinfeksjoner, lungebetennelse, hjernehinnebetennelse (meningitt) og blodforgiftning, en systematisk inflammatorisk respons som forårsaker sjokk, feber og en massiv vasodilatasjon og død.[64] Likevel er disse organismene en del av normalfloraen og lever vanligvis på huden eller i nesen uten å gi årsak til sykdom. Andre organismer forårsaker alltid sykdommer hos mennesket, slik som Rickettsia, som infiserer cellen og kan kun vokse og reprodusere seg innen celler hos andre organismer. En art av Rickettsia forårsaker tyfoidfeber, mens andre er årsak til flekkfeber (eng, «Rocky Mountain spotted fever»). Klamydia, en annen rekke som infiserer andre celler, inneholder arter som er årsak til lungebetennelse, urinveisinfeksjon og kan være en årsak til koronar hjertesykdom.[65] Noen bakterier som Pseudomonas aeruginosa, Burkholderia cenocepacia, og Mycobacterium avium er opportunistiske patogener, som betyr at de forårsaker sykdom kun når immunforsvaret er svekket eller ved cystisk fibrose.[66][67]
Bakterielle infeksjoner kan bli behandlet med antibiotika, som er klassifisert som baktericid når de dreper bakterier, eller bakteriostatisk hvis de forhindrer vekst. De ulike typene antibiotika forhindrer forskjellige mekanismer hos de ulike patogene bakteriene. Et eksempel på hvordan antibiotika produserer selektiv toksisitet er kloramfenikol og puromycin, som forhindrer bakterielle ribosomers arbeid, men påvirker ikke det strukturelt ulike eukaryotiske ribosomet.[68] Antibiotika blir benyttet til å behandle sykdom både hos mennesker og dyr. Overforbruk av antibiotika er en årsak til at bakterier blir resistente overfor det. Infeksjoner forhindres ved å bruke antiseptiske metoder som sterilisering av huden ved bruk av sprøyter og til å rense sår. Kirurgisk- og tannlegeutstyr blir også sterilisert for å unngå forurensing og infeksjon av bakterier. Desinfeksjon av overflater blir benyttet til å drepe bakterier og andre patogener for å redusere risikoen for infeksjon.
Bakterier i industri[rediger | rediger kilde]
Bakterier, ofte Lactobacillus i kombinasjon med mugg- og gjærsopp har blitt brukt i over tusen år til fremstilling av gjæret mat som ost, soyasaus, surkål, eddik og yoghurt.[69][70] Bakterier blir brukt til å bryte ned avfall på grunn av deres ferdighet til å bryte ned en stor variasjon av organisk materiale. Bakteriene har også den muligheten at de kan bryte ned hydrokarboner i petroleumsprodukter og blir ofte benyttet til å rydde opp i oljesøl.[71] Kunstgjødsel ble brukt på strender i Prince William-sundet i et forsøk på å påskynde vekst av en naturlig voksende bakterie etter det beryktede Exxon Valdez-oljesølet i 1989. Dette var effektivt på de strendene som ikke var for tilsølet med olje. Bakterier blir også brukt til å bryte ned andre farlige kjemikalier.[72] De er også viktig innen kjemisk industri i produksjon av rene kjemikalier[73] som blir brukt innen legemidler eller kjemikalier brukt i landbruket.[74]
Ofte blir bakterier benyttet i pesticid i biologisk kontroll av skadedyr. Bacillus thuringiensis (også kalt BT) er mest vanlig, som er en Gram-positive bakterie som er vanlig i jord. Delarterr av denne bakterien blir brukt som et sommerfugl-spesifiskt insektmiddel under forskjellige varemerker som for eksempel Dipel og Thuricide.[75] På grunn av deres spesifikke egenskap, er disse pesticidene ikke sett på som skadelig på miljøet og med liten eller ingen effekt på mennesker, dyr eller fordelaktige insekter.[76][77]
Bakteriene er arbeidshestene i områdene mikrobiologi, genetikk og biokjemi på grunn av deres enkle oppbygning og evnen til å vokse raskt. Ved å lage mutasjoner i bakterienes DNA og ved å undersøke de resulterende fenotyper, kan forskere avgjøre funksjoner av gener, enzymer og metabolske gangstier, og for så å anvende denne kunnskapen på høyere organismer.[78]
Gramfarging[rediger | rediger kilde]

Utdypende artikkel: Gramfarging
En annen inndeling av bakteriene er mellom grampositive og gramnegative bakterier. Denne baserer seg på en fargeteknikk som kalles gramfarging og som indikerer forskjeller i celleveggens oppbygning. Denne metoden ble oppkalt etter dansken Hans Christian Gram. Gram oppfant metoden i slutten av forrige århundre der han jobbet med å utvikle en prosedyre for å identifisere bakterieinfiserte dyreceller. Gramfarging utføres i flere trinn med forskjellige fargevæsker. Etter prosedyren blir bakteriene enten røde eller blå. Grampositive blir blåfiolette og gramnegative blir røde/rosa.[79]
Se også[rediger | rediger kilde]
Fotnoter og referanser[rediger | rediger kilde]
- ^ Bess B. Ward, «How many species of prokaryotes are there?»
- ^ Vanskelig å sette et estimat på antall arter prokaryoter
- ^ Fredrickson J, Zachara J, Balkwill D; m.fl. (2004). «Geomicrobiology of high-level nuclear waste-contaminated vadose sediments at the hanford site, Washington state». Appl Environ Microbiol. 70 (7): 4230–41. PMID 15240306. Arkivert fra originalen 29. september 2008. Besøkt 5. august 2007.
- ^ Whitman W, Coleman D, Wiebe W (1998). «Prokaryotes: the unseen majority». Proc Natl Acad Sci U S A. 95 (12): 6578–83. PMID 9618454. Arkivert fra originalen 5. mars 2008. Besøkt 5. august 2007.
- ^ Rappé M, Giovannoni S. «The uncultured microbial majority». Annu Rev Microbiol. 57: 369–94. PMID 14527284.
- ^ Sender, R., Fuchs, S. & Milo, R. (2016). «Revised estimates for the number of human and bacteria cells in the body» (PDF). BioReXiv beta. Besøkt 14. januar 2016.
- ^ Caplex, «Bakterier»
- ^ 2002 WHO mortality data Accessed 20 January 2007
- ^ Ishige T, Honda K, Shimizu S (2005). «Whole organism biocatalysis». Curr Opin Chem Biol. 9 (2): 174–80. PMID 15811802.
- ^ Dyreceller=Våre celler, inneholder cellekjerne. Se Eukaryoter.
- ^ Woese C, Kandler O, Wheelis M (1990). «Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya». Proc Natl Acad Sci U S A. 87 (12): 4576–9. PMID 2112744.
- ^ Hallingbäck, T., et al: Nationalnyckeln till Sveriges flora och fauna, «Bladmossor», Artsdatabanken, SLU Uppsala, 2006, side 14-16.
- ^ Nature – Francesca D. Ciccarelli, Tobias Doerks, Christian von Mering, Christopher J. Creevey, Berend Snel,Peer Bork: «Toward Automatic Reconstruction of a Highly Resolved Tree of Life», Nature Nr 311, 2006. De kaller arkebakterier for en fyle («Phylum»).
- ^ Hallingbäck, T., et al: Nationalnyckeln till Sveriges flora och fauna, «Bladmossor», Artsdatabanken, SLU Uppsala, 2006.
- ^ Nature – Francesca D. Ciccarelli, Tobias Doerks, Christian von Mering, Christopher J. Creevey, Berend Snel,Peer Bork: «Toward Automatic Reconstruction of a Highly Resolved Tree of Life», Nature Nr 311, 2006.
- ^ Encyclopædia Britannica – agar
- ^ Encyclopædia Britannica, Inc., «Evolution of bacteria»
- ^ DeLong E, Pace N (2001). "Environmental diversity of bacteria and archaea". Syst Biol 50 (4): 470–78. PMID .
- ^ Brown J, Doolittle W (1997). "Archaea and the prokaryote-to-eukaryote transition". Microbiol Mol Biol Rev 61 (4): 456–502. PMID
- ^ a b c Espen Aas (2017) – Vi har funnet verdens eldste form for liv. Urix (web), NRK, 1. mars 2017. Besøkt 2017-03-01
- ^ Nano, 1 milliarddel. Se nano
- ^ Mississippi State University – Nannobacteria Arkivert 7. januar 2006 hos Wayback Machine.
- ^ a b Biosite - Gram negative og gram positive bakterier
- ^ a b c d e f g h * Michael T. Madigan, John M. Martinko, Jack Parker (2003). Brock Biology of Microorganisms, Tenth Edition. Pearson Education, Inc.
- ^ a b c The Bacterial Cell Wall[død lenke]
- ^ a b Medical Microbiology - A Brief Introduction Arkivert 2. juni 2007 hos Wayback Machine.
- ^ «Cyberlipid Center-Resource site for lipid studies - Lipopolysaccharides». Arkivert fra originalen 2. juni 2007. Besøkt 31. mai 2007.
- ^ Berg J., Tymoczko J. and Stryer L. (2002) Biochemistry. W. H. Freeman and Company ISBN
- ^ Poehlsgaard J, Douthwaite S (2005). "The bacterial ribosome as a target for antibiotics". Nat Rev Microbiol 3 (11): 870–81. PMID
- ^ a b Microbiology and Bacteriology - 2-34 Endospores are very resistant structures
- ^ Prokaryotic cell structure: Endospores Arkivert 8. juli 2007 hos Wayback Machine.
- ^ Nealson K (1999). "Post-Viking microbiology: new approaches, new data, new insights". Orig Life Evol Biosph 29 (1): 73–93. PMID 11536899
- ^ a b UiO Biologi - Regnum Bacteria - Bakterieriket
- ^ Stephen T. Abedon, Ohio State University - Important words and concepts from Chapter 5, Black, 1999 (3/28/2003) Arkivert 11. mai 2007 hos Wayback Machine.
- ^ Microbiology and Bacteriology - Macronutrients, Micronutrients and Growth Factors
- ^ Michael Knee, The Ohio State University – Bacteria Arkivert 15. juni 2007 hos Wayback Machine.
- ^ Sulfid er en fellesbetegnelse for en rekke stoffer
- ^ Polymerer betyr: å lage mange
- ^ For eksempel en kuleform har areal og volum , slik at
- ^ Nakabachi A, Yamashita A, Toh H, Ishikawa H, Dunbar H, Moran N, Hattori M (2006). «The 160-kilobase genome of the bacterial endosymbiont Carsonella». Science. 314 (5797): 267. PMID 17038615.
- ^ Pradella S, Hans A, Spröer C, Reichenbach H, Gerth K, Beyer S (2002). «Characterisation, genome size and genetic manipulation of the myxobacterium Sorangium cellulosum So ce56». Arch Microbiol. 178 (6): 484-92. PMID 12420170.
- ^ Helsetilsynet.no, «BORRELIOSE (LYMES SYKDOM) Infeksjon med Borrelia burgdorferi (...)»[død lenke]
- ^ Hinnebusch J, Tilly K (1993). «Linear plasmids and chromosomes in bacteria». Mol Microbiol. 10 (5): 917-22. PMID 7934868.
- ^ Hastings P, Rosenberg S, Slack A (2004). «Antibiotic-induced lateral transfer of antibiotic resistance». Trends Microbiol. 12 (9): 401–4. PMID 15337159.
- ^ Brüssow H, Canchaya C, Hardt W (2004). «Phages and the evolution of bacterial pathogens: from genomic rearrangements to lysogenic conversion». Microbiol Mol Biol Rev. 68 (3): 560–602. PMID 15353570.
- ^ Denamur E, Matic I (2006). «Evolution of mutation rates in bacteria». Mol Microbiol. 60 (4): 820–7. PMID 16677295.
- ^ Wright B (2004). «Stress-directed adaptive mutations and evolution». Mol Microbiol. 52 (3): 643–50. PMID 15101972.
- ^ Bokmålsordboka, «konjugere»
- ^ Davison J (1999). «Genetic exchange between bacteria in the environment». Plasmid. 42 (2): 73–91. PMID 10489325.
- ^ MicrobiologyBytes, Bacterial Motility Arkivert 4. august 2007 hos Wayback Machine.
- ^ a b Randall Institute, King's College London, UK., The bacterial flagella motor.
- ^ Med flagellisjon menes hvordan flagellene er arrangert
- ^ ISCID Encyclopedia of Science and Philosophy - Flagellin
- ^ «Flagellin - definition from Biology-Online.org». Arkivert fra originalen 27. april 2007. Besøkt 31. mai 2007.
- ^ ^ a b c Bardy S, Ng S, Jarrell K (2003). "Prokaryotic motility structures". Microbiology 149 (Pt 2): 295–304. PMID 12624192.
- ^ Engelsk: Interspecies hydrogen transfer
- ^ Stams A, de Bok F, Plugge C, van Eekert M, Dolfing J, Schraa G (2006). "Exocellular electron transfer in anaerobic microbial communities". Environ Microbiol 8 (3): 371–82. PMID 16478444
- ^ Barea J, Pozo M, Azcón R, Azcón-Aguilar C (2005). "Microbial co-operation in the rhizosphere". J Exp Bot 56 (417): 1761–78. PMID 15911555
- ^ O'Hara A, Shanahan F (2006). "The gut flora as a forgotten organ". EMBO Rep 7 (7): 688–93. PMID 16819463.
- ^ Zoetendal E, Vaughan E, de Vos W (2006). "A microbial world within us". Mol Microbiol 59 (6): 1639–50. PMID 16553872.
- ^ Gorbach S (1990). "Lactic acid bacteria and human health". Ann Med 22 (1): 37–41. PMID 2109988
- ^ Eksempel på probiotika i Norge: Idoform
- ^ Salminen S, Gueimonde M, Isolauri E (2005). "Probiotics that modify disease risk". J Nutr 135 (5): 1294–8. PMID 15867327
- ^ Fish D. "Optimal antimicrobial therapy for sepsis". Am J Health Syst Pharm 59 Suppl 1: S13–9. PMID 11885408
- ^ Belland R, Ouellette S, Gieffers J, Byrne G (2004). "Chlamydia pneumoniae and atherosclerosis". Cell Microbiol 6 (2): 117–27. PMID 14706098
- ^ Heise E. "Diseases associated with immunosuppression". Environ Health Perspect 43: 9–19. PMID 7037390.
- ^ Saiman, L. "Microbiology of early CF lung disease". Paediatr Respir Rev.volume=5 Suppl A: S367–369. PMID 14980298
- ^ Yonath A, Bashan A (2004). "Ribosomal crystallography: initiation, peptide bond formation, and amino acid polymerization are hampered by antibiotics". Annu Rev Microbiol 58: 233–51. PMID 15487937
- ^ Johnson M, Lucey J (2006). "Major technological advances and trends in cheese". J Dairy Sci 89 (4): 1174–8. PMID 16537950
- ^ Hagedorn S, Kaphammer B (1994). "Microbial biocatalysis in the generation of flavor and fragrance chemicals". Annu. Rev. Microbiol. 48: 773-800. doi:10.1146/annurev.mi.48.100194.004013. PMID 7826026
- ^ Cohen Y (2002). "Bioremediation of oil by marine microbial mats". Int Microbiol 5 (4): 189–93. PMID 12497184
- ^ Neves LC, Miyamura TT, Moraes DA, Penna TC, Converti A (2006). "Biofiltration methods for the removal of phenolic residues". Appl. Biochem. Biotechnol. 129-132: 130-52. PMID 16915636
- ^ I syntetisering av kjemikalier dannes biprodukter. Eksempel, forskjellige stereoisomeri som vil ha andre egenskaper.
- ^ Liese A, Filho M (1999). "Production of fine chemicals using biocatalysis". Curr Opin Biotechnol 10 (6): 595–603. PMID 10600695
- ^ Aronson AI, Shai Y (2001). "Why Bacillus thuringiensis insecticidal toxins are so effective: unique features of their mode of action". FEMS Microbiol. Lett. 195 (1): 1-8. PMID 11166987
- ^ Bozsik A (2006). "Susceptibility of adult Coccinella septempunctata (Coleoptera: Coccinellidae) to insecticides with different modes of action". Pest Manag Sci 62 (7): 651–4. PMID 16649191
- ^ Chattopadhyay A, Bhatnagar N, Bhatnagar R (2004). "Bacterial insecticidal toxins". Crit Rev Microbiol 30 (1): 33–54. PMID 15116762
- ^ Serres M, Gopal S, Nahum L, Liang P, Gaasterland T, Riley M (2001). "A functional update of the Escherichia coli K-12 genome". Genome Biol 2 (9): REEARCH0035. PMID 11574054
- ^ University of Pennsylvania Health System - Gram stain history and mechanism



Litteratur[rediger | rediger kilde]
Norsk[rediger | rediger kilde]
- Tjade, Trygve (2002). Medisinsk mikrobiologi og infeksjonssykdommer. Fagbokforlaget.
- Jessica Lönn-Stensrud (2019) Vi må snakke om bakterier. Dine viktigste venner ISBN 9788248924258
- Jessica Lönn-Stensrud (2016) Bakterienes forunderlige verden ISBN 9788215024967
Engelsk[rediger | rediger kilde]
- Alcamo, I. Edward. Fundamentals of Microbiology. 6th ed. Menlo Park, California: Benjamin Cumming, 2001. ISBN
- Atlas, Ronald M. Principles of Microbiology. St. Louis, Missouri: Mosby, 1995. ISBN
- Michael T. Madigan, John M. Martinko, Jack Parker (2003). Brock Biology of Microorganisms, Tenth Edition. Pearson Education, Inc.
- Glazer, A.N. & Nikaido, H. (1995) Microbial biotechnology. Fundamentals of Applied Microbiology. W.H.Freeman & Company.
- Holt, John. G. Bergey's Manual of Determinative Bacteriology. 9th ed. Baltimore, Maryland: Williams and Wilkins, 1994.
- Hugenholtz P, Goebel BM, Pace NR (1998). «Impact of culture-independent studies on the emerging phylogenetic view of bacterial diversity». J Bacteriol. 180 (18): 4765–74. PMID.
- Madigan, Michael and Martinko, John. Brock Biology of Microorganisms. 11th ed. Prentice Hall, 2005. ISBN
- Tortora, Gerard; Funke, Berdell; Case, Christine. Microbiology: An Introduction. 8th ed. Benjamin Cummings, 2003.
- Witzany G. "Bio-Communication of Bacteria and their Evolutionary Roots in Natural Genome Editing Competences of Viruses." Open Evolution Journal 2: 44-54, 2008.
Eksterne lenker[rediger | rediger kilde]
- (en) bakterier – oversikt og omtale av artene i WORMS-databasen
- (en) bakterier i Encyclopedia of Life
- (en) bakterier i Global Biodiversity Information Facility
- (no) bakterier hos Artsdatabanken
- (sv) bakterier hos Dyntaxa
- (en) bakterier hos ITIS
- (en) bakterier hos MycoBank
- (en) bakterier hos NCBI
- (en) Kategori:Bacteria – bilder, video eller lyd på Wikimedia Commons
- Bacteria – detaljert informasjon på Wikispecies
- Nettdoktor.no – «Helseråd om virus og bakterier»
- UiO Biologi – «Bakterier»
- UMB – «Bakterier – våre gode hjelpere»
- Universitetet i Tromsø, Institutt for biologi, «mer om metabolisme»
- UiO Biologi – Evolusjon
- Tidsskrift for Den norske lægeforening - «Hva gjør bakterier patogene?»
- «Bakterier og virus» av Halvor Aarnes (2003) (PDF-fil)
- Hva er bakterier? - artikkel fra forskning.no 12.7.02
- (engelsk) Bacteriamuseum Arkivert 10. juni 2007 hos Wayback Machine.
- (engelsk) Biology of Microorganisms, Eighth Edition. «On-line study guide»
- (engelsk) Brock Biology of Microorganisms, 10th edition. «Animated Tutorials»
- (engelsk) «Bacterial Nomenclature Up-To-Date» fra DSMZ Arkivert 29. september 2007 hos Wayback Machine.
- (engelsk) Video Arkivert 5. november 2015 hos Wayback Machine. av bakterier «svømmer» og av deres bevegelse m.m.
- (engelsk) «Planet of the Bacteria» av Stephen Jay Gould
- (engelsk) «The largest bacteria» Arkivert 11. desember 2012 hos Wayback Machine.
- Høgskolen i Oslo, Laboratoriekurs, modul 1, mikrobiologi